|
-------In
the Patagonian steppe, vegetation is composed
of shrubs and tussock grasses. They are arranged in two patch types:
shrubs encircled by a dense ring of tussocks grasses and tussock grasses
interspersed with bare soil (Soriano et al. 1994). A model has
been 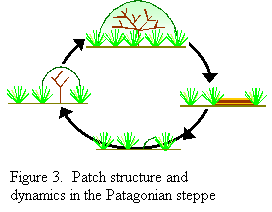 proposed
for the origin and maintenance of this patchy structure in which spatial
heterogeneity is viewed as reflecting phases of a cyclical succession
(Fig. 3). When a shrub seedling establishes, it grows, and after achieving
a certain size generates favorable conditions for grass establishment.
These favorable conditions result in the construction of a dense ring
of grasses surrounding the shrub. The building of the ring represents
the construction phase of the cycle. When the shrub dies and aerial
protection disappears, the density of grasses becomes higher than the
current carrying capacity of the ring that results in the death of some
tussocks. The ring disappears as an entity, and grasses appear interspersed
with bare soil patches forming the other patch type of the Patagonian
steppe. This would be the destruction phase of the cycle. proposed
for the origin and maintenance of this patchy structure in which spatial
heterogeneity is viewed as reflecting phases of a cyclical succession
(Fig. 3). When a shrub seedling establishes, it grows, and after achieving
a certain size generates favorable conditions for grass establishment.
These favorable conditions result in the construction of a dense ring
of grasses surrounding the shrub. The building of the ring represents
the construction phase of the cycle. When the shrub dies and aerial
protection disappears, the density of grasses becomes higher than the
current carrying capacity of the ring that results in the death of some
tussocks. The ring disappears as an entity, and grasses appear interspersed
with bare soil patches forming the other patch type of the Patagonian
steppe. This would be the destruction phase of the cycle.
A heterogeneous vegetation structure results in the
concentration of resources underneath individual plants, patches, or
stripes. Two decades ago, Charley and West, (1975)
introduced the term islands of fertility to describe the accumulation
of nutrients underneath the canopy of shrubs in the semi-desert of Utah.
Similarly, in the Chilean matorral, the concentration of soil nitrogen,
phosphorus, and soil organic matter were higher underneath than outside
the canopies of shrubs (Gutierrez
et al. 1993). Smaller individuals such as those of the grass
species Bouteloua gracilis
were able to significantly modify the concentration of nitrogen and
carbon underneath their canopies (Hook
et al. 1991). At a coarser scale, the upper soil layer of the
mulga groves had higher available nitrogen than the inter-groves (Ludwig and Tongway 1995). Phosphorus
being an element less mobile in the soil showed similar trends but only
in the uppermost layer.
What is the effect of this heterogeneous distribution
of resources on the functioning of the entire ecosystem? Does it enhance
or reduce production? Does it enhance or reduce ecosystem functioning
in general? The answers to these questions have profound theoretical
and applied implications. They can influence our understanding of the
functioning of arid rangelands and our selection of management techniques
that may try to either avoid or take advantage of heterogeneity. Schlesinger,
et al. (1990) suggested that heterogeneity
in the distribution of resources was a result of desertification while
others indicate that is necessary for the survival of many plants and
animals in arid and semiarid rangelands (Stafford
Smith and Pickup 1990).
|
|
-------We
suggest that the model that relates production and precipitation is
linear and has two thresholds (Fig. 4A). Below a minimum precipitation
threshold production is zero, and beyond a maximum precipitation threshold
further increases in precipitation do not result in increases in production.
There is empirical support of the existence of the minimum and maximum
thresholds. Analysis of the production patterns of 9498 grassland sites
in the Central Grassland Region of North America indicated that primary
production and precipitation are positively correlated and the linear
model has a negative y-intercept (Sala
et al. 1988):
ANPP=-34 + 0.6*APPT
where
ANPP is aboveground net primary production and APPT is annual precipitation.
This equation can be rearranged as:
ANPP= 0.6*(APPT-56)
where
0.6 represents the average water use efficiency and 56 is the ineffective
precipitation or zero-yield intercept (Noy-Meir
1973). The analysis of the production patterns of forests, grasslands,
and deserts suggested the existence of an upper threshold in the response
of production to precipitation (Webb
et al. 1978, Webb et al. 1983).
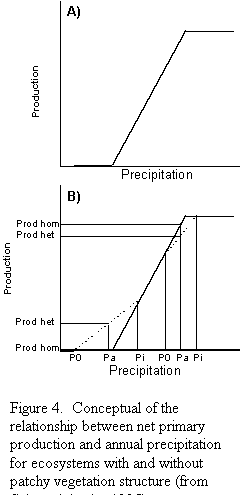 This model has important implications for the effect of spatial
heterogeneity on total ecosystem production based on the idea that in
a patchy environment there are areas which concentrate resources, sinks,
and areas which provide resources, sources. In the case of water, those
are run-on and run-off areas. The amount of water redistributed depends
on the proportion of sources and sinks, and on the fraction of precipitation
which runs off which in turn depends on the magnitude of the precipitation
event, the soil water content at the time
of the event, topography, plant cover, and stocking rate (Branson
et al. 1981). If average
precipitation falls below the lower threshold and
it is homogeneously distributed, available water is (Pa) and production
should be zero (Fig. 4B). If there is redistribution, the source areas
will give up some water and will end up with less available water (Po),
and production will continue to be zero. However, the sink areas will
receive precipitation plus run-on that will result in a higher water
availability (Pi) which now is higher than the threshold. Consequently,
there will be production in the sink areas. This analysis suggests that
in arid regions heterogeneous systems may have higher production than
homogeneous systems. On the contrary, in mesic grasslands in which average
precipitation is close to the upper threshold, heterogeneous systems
should have lower production than homogeneous systems. This analysis
was performed for water redistribution and it can be similarly performed
for some other resources that limit production at different points in
time. Nitrogen and phosphorus can be redistributed as water. In the
case of nutrients, biological mechanisms can be invoked in addition
to abiotic mechanisms to account for the redistribution of nutrients.
Roots forage for nutrients beyond canopy projection and most of the
aboveground litter is deposited underneath the canopy. Finally, animals
may contribute to the redistribution of nutrients and concentration
in patches.
This model has important implications for the effect of spatial
heterogeneity on total ecosystem production based on the idea that in
a patchy environment there are areas which concentrate resources, sinks,
and areas which provide resources, sources. In the case of water, those
are run-on and run-off areas. The amount of water redistributed depends
on the proportion of sources and sinks, and on the fraction of precipitation
which runs off which in turn depends on the magnitude of the precipitation
event, the soil water content at the time
of the event, topography, plant cover, and stocking rate (Branson
et al. 1981). If average
precipitation falls below the lower threshold and
it is homogeneously distributed, available water is (Pa) and production
should be zero (Fig. 4B). If there is redistribution, the source areas
will give up some water and will end up with less available water (Po),
and production will continue to be zero. However, the sink areas will
receive precipitation plus run-on that will result in a higher water
availability (Pi) which now is higher than the threshold. Consequently,
there will be production in the sink areas. This analysis suggests that
in arid regions heterogeneous systems may have higher production than
homogeneous systems. On the contrary, in mesic grasslands in which average
precipitation is close to the upper threshold, heterogeneous systems
should have lower production than homogeneous systems. This analysis
was performed for water redistribution and it can be similarly performed
for some other resources that limit production at different points in
time. Nitrogen and phosphorus can be redistributed as water. In the
case of nutrients, biological mechanisms can be invoked in addition
to abiotic mechanisms to account for the redistribution of nutrients.
Roots forage for nutrients beyond canopy projection and most of the
aboveground litter is deposited underneath the canopy. Finally, animals
may contribute to the redistribution of nutrients and concentration
in patches.
In this project, we address
both the effect of land-use change (i.e., grazing) and the effect of
climate change (i.e., precipitation) on the spatial pattern of arid
ecosystems. Our main underlying objective is to identify how these two
driving forces control the vegetation spatial patterns and how they
affect the functioning of ecosystems. We approached this issue with
a combination of observation and field manipulative experiments.
|
|
-----
Referencesefer
- Branson, F.,
G. Gifford, K. Renard, and R. Hadley. 1981. Runoff and water harvesting.
Pages 73-110 in E. H. Reid,
editor. Rangeland Hydrology. Kendall/Hunt Publishing Company, Dubuque,
Iowa.
- Charley, J.,
and N. West. 1975. Plant-induced soil chemical patterns in some shrub-dominated
semi-desert ecosystems in Utah. Journal of Ecology 63:945-963.
- Gutierrez, J., P. Meserve, L. Contreras,
H. Vasquez, and F. Jaksic. 1993. Spatial distribution of soil
nutrients and ephemeral plants underneath and outside the canopy of
Porlieria chilensis shrubs (Zygophyllaceae) in arid coastal Chile.
Oecologia 95:347-352.
- Hook, P., I.
Burke, and W. Lauenroth. 1991. Heterogeneity of soil and plant N and
C associated with individual openings in North American shortgrass
steppe. Plant and Soil 138:247-256.
- Ludwig, J.,
and D. Tongway. 1995. Spatial organisation of landscapes and its function
in semi-arid woodlands, Australia. Landscape Ecology 10:51-63.
- Noy-Meir, I.
1973. Desert ecosystems: environment and producers. Ann. Rev. Ecol.
Syst. 4:25-52.
- Sala, O. E.,
W. J. Parton, W. K. Lauenroth, and L. A. Joyce. 1988. Primary production
of the central grassland region of the United States. Ecology. 69:40-45.
- Schlesinger,
W. H., J. F. Reynolds, G. L. Cunningham, L. F. Huenneke, W. M. Jarrell,
R. A. Virginia, and W. G. Whitford. 1990. Biological feedbacks in
global desertification. Science 247:1043-1048.
- Soriano, A.,
O. E. Sala, and S. B. Perelman. 1994. Patch structure and dynamics
in a Patagonian arid steppe. Vegetatio. 111:127-135.
- Stafford Smith,
D., and G. Pickup. 1990. Pattern and production in arid lands. Proceedings
Ecological Society of Australia 16:195-200.
- Webb, W., S.
Szarek, W. K. Lauenroth, R. Kinerson, and M. Smith. 1978. Primary
productivity and water use in native forest, grassland, and desert
ecosystems. Ecology. 59:1239-1247.
- Webb, W. L.,
W. K. Lauenroth, and S. R. Szarell. 1983. Primary production and abiotic
controls in forests, grasslands and desert ecosystems in the United
States. Ecology. 64:134-151.
|