Nos presentamos.
Revisamos algunos de los conocimientos de matemática, física y biología elementales para poder aprovechar el curso de Ecología de los Ecosistemas.
Planteamos el objetivo central de este curso que es comprender los mecanismos básicos de la productividad de los agroecosistemas. En relación con esta cuestión distinguimos entre los productos vegetales y los productos animales, vimos que los productos animales derivan de productos vegetales y que los productos vegetales, a su vez, son construidos en el agroecosistema, principalmente a partir de dióxido de carbono de la atmósfera, agua y luz. Mencionamos que para comprender estos mecanismos vamos a necesitar examinar aspectos de la estructura y del funcionamiento de los agroecosistemas.
Observamos la morfología de diferentes especies de plantas superiores y ejercitamos la identificación de los tres tipos de órganos: hojas, tallos y raíces.
Describimos la anatomía de las hojas y de las raíces poniendo especial atención a la manera en que esa anatomía se adecua para la absorción de luz y para el intercambio de agua y gases (como el dióxido de carbono por ejemplo) entre las plantas y el medio que las rodea. Para cada órgano examinamos las vías de entrada y de salida de agua y de carbono. No pusimos especial énfasis en las entradas y salidas de oxígeno. Por eso les recomiendo que, como ejercicio, analicen cómo entra y cómo sale el oxígeno de los diferentes órganos de las plantas.
Propusimos que la tasa de intercambio de energía y substancias entre las plantas y el medio depende de: (1) la superficie del área de contacto, (2) su permeabilidad (o la resistencia que es su inversa) y (3) el gradiente de concentración de la substancia. En particular analizamos cómo varían estos factores en relación con la entrada de agua en las raíces y con la entrada de dióxido de carbono y la salida de agua de las hojas. Vimos que el mecanismo de apertura y cierre de los estomas hace que la entrada de dióxido de carbono dependa del estado de hidratación de las hojas.
Examinamos con cierto detalle el proceso de la fotosíntesis.
Durante la discusión vimos primero qué es la fotosíntesis en términos generales y en qué se diferencia de la respiración. Como otros procesos de asimilación, la fotosíntesis requiere energía. En particular utiliza energía lumínica.
Luego examinamos rápidamente el "aparato" que produce la fotosíntesis en las escalas de órgano, tejido, célula y cloroplasto con el fin de entender cómo estas estructuras resultan adecuadas para esa función.
Después examinamos en mayor detalle cómo ocurre la fotosíntesis en las plantas C3. Diferenciamos las etapas fotoquímica y bioquímica (o de fijación de carbono) en términos de los lugares donde ocurren (tilacoide y estroma), de sus principales agentes (clorofila/pigmentos y rubisco) y de sus productos de entrada y salida (agua, luz, NADP, ADP, etc.). Este análisis nos permitió comenzar a pensar en los factores que controlan la tasa de fotosíntesis. El miércoles próximo vamos a volver a ver esta cuestión en más detalle porque es fundamental para entender como se puede regular la productividad de los cultivos.
A partir del análisis del mecanismo de fotosíntesis de las plantas C3, presentamos el de las plantas C4 que simplemente consiste en el agregado de un paso previo en la etapa bioquímica que, si bien consume un poco de energía asegura que la concentración de dióxido de carbono en los cloroplastos sea normalmente alta.
Definimos a la fotosíntesis neta, comenzamos a examinar las unidades adecuadas para medirla y definimos el punto de compensación.
Finalmente los alumnos trabajaron en grupos con los problemas 1 a 4.
Discutimos los problemas 1 a 4. Mediante esta discusión continuamos con nuestro análisis de la fotosíntesis, el proceso por el cual las plantas asimilan el carbono. Volvimos a identificar las diferentes etapas que este proceso tiene en las plantas C3 y C4 y vimos que la fotolisis del agua depende de la incidencia de luz y la fijación de CO2 (por carboxilación del difosfato de ribulosa RudP) depende justamente de la presencia de CO2 en los cloroplastos. También vimos que la presencia de CO2 en los cloroplastos depende en las plantas C3 de la difusión y en las plantas C4 de la llegada de ácido málico desde las células del mesófilo donde ocurre la carboxilación del fosfoenol-piruvato (PEP).
Este análisis, nos introdujo en el concepto de FACTOR LIMITANTE. Dijimos que, por ejemplo, la intensidad de luz es un factor limitante para la fotosíntesis si cuando se aumenta la intensidad de luz se obtiene como respuesta un aumento en la tasa de fotosíntesis. En particular, vimos que la intensidad de luz es un factor limitante de la tasa de fotosíntesis siempre que está por debajo del PUNTO DE SATURACION.
Tomando primero la intensidad luz y luego la concentración de CO2 como ejemplos, vimos cómo son las curvas de respuesta de la fotosíntesis a factores que pueden volverse limitantes. En relación con estas curvas nos tomamos bastante tiempo para aprender a definir los PUNTOS DE COMPENSACION y de SATURACION.
Además nos concentramos en algunos aspectos de la luz y del CO2 como controles de la fotosíntesis. En relación con la luz: discutimos las unidades apropiadas para medir la intensidad de luz, vimos cómo se reparte la luz que incide sobre una hoja, vimos cómo la intensidad de luz que llega a una hoja depende de su inclinación y vimos que la cantidad de luz absorbida por un cultivo depende del Índice de Área Foliar (la superficie de hojas que hay por cada unidad de superficie de suelo). En relación con el CO2, vimos las diferencias en la respuesta de las plantas C3 y C4 a su concentración.
Dedicamos la mayor parte de la clase a discutir el problema 5. Sobre la base de este problema examinamos en detalle la influencia de los principales factores limitantes de la tasa de fotosíntesis neta de una hoja:
- la intensidad de luz
- la concentración de dióxido de carbono
- la disponibilidad de agua
- la disponibilidad de nitrógeno
- la temperatura
Además ejercitamos la habilidad para realizar cálculos que involucran cantidades de energía y de materia. Mediante los cálculos que realizamos, pudimos comparar la eficiencia con que aprovechan la luz incidente las hojas de una planta C3 sometidas diferente intensidad de luz. Encontramos que, según los datos de que disponíamos, bajo una intensidad mediana (100Watt/m²) la cantidad de energía incidente necesaria para fijar 1 mg de glucosa es mucho menor bajo una intensidad de luz alta (300Watt/m²).Finalmente notamos que una manera en que las plantas pueden mejorar el aprovechamiento de la luz incidente es inclinar las hojas y aumentar el Índice de Área Foliar.
En el tiempo que nos quedaba comenzamos a analizar la dinámica del agua en el sistema suelo-planta-atmósfera. En relación con este tema:
- Identificamos al déficit hídrico como el factor que con más frecuencia limita en efecto a la
fotosíntesis (y como resultado al crecimiento de las plantas).
- Vimos que la transpiración tiene 2 consecuencias principales: (1) mantiene las hojas frescas y
(2) causa la absorción de agua del suelo.
- Vimos que, en el suelo, el
agua está en poros de diferentes tamaños y que la fuerza con la cual está
retenida es mayor cuanto menor es el diámetro de los poros.
- Este mismo principio se aplica
al agua retenida en las paredes de las células del mesófilo de las hojas que
están en contacto con espacio intercelular hacia los cuales el agua se evapora.
Allí, los poros o conductos donde está el agua son los que separan a las micro
fibrillas de celulosa y son extremadamente delgados; por eso retienen el agua
con una fuerza muy grande.
- Esta fuerza se transmite, al evaporarse el agua, como una tensión que arrastra
el agua hacia arriba desde el suelo a lo largo de las raíces y los tallos gracias a la
cohesión existente entre las moléculas de agua.
Más o menos en este punto
estábamos cuando se nos acabó el tiempo disponible para la clase. Por eso no
alcanzamos a formalizar el concepto de POTENCIAL HÍDRICO, que es la energía
con la cual el agua está retenida (por ejemplo en el suelo o en el mesófilo de
una hoja) y que se mide en unidades de presión como por ejemplo atmósferas o
Megapascales (10 atm = 1MPa). Este concepto es necesario para resolver el
problema 2 de la Evaluación Orientativa. Les sugiero que, aunque aún no lo
discutimos en clase, intenten igual resolver este problema ya que les servirá
de preparación para terminar de discutir el tema el miércoles próximo. Tengan en
cuenta que cuánto menor es el valor del Potencial Hídrico más fuertemente está retenida el agua.
Por eso, un suelo con Potencial Hídrico de –0,3MPa retiene el agua con poca
fuerza que uno con Potencial Hídrico de –1,2MPa la retiene con mucha fuerza. Las plantas cultivadas
pueden mantener los estomas abiertos cuando el Potencial Hídrico del suelo es
de –0,3MPa y en cambio cuando es tan bajo como 1,2MPa los cierran.
Discutimos en detalle el proceso del movimiento del agua desde el suelo hasta la atmósfera a través de las plantas y revisamos, de paso, conceptos básicos de ciencia del suelo.
Para organizar la discusión propusimos examinar las siguientes cuestiones: ¿dónde ocurre el proceso? ¿cómo ocurre? ¿en qué unidades se miden las magnitudes involucradas en el proceso? y ¿qué factores regulan el proceso y cómo?
En relación con la primera cuestión (¿dónde?), identificamos al suelo, las plantas y la atmósfera como los componentes principales del sistema en donde ocurre el movimiento del agua que nos interesaba analizar. Además recordamos la estructura de las plantas, compuestas por raices, tallos y hojas, a través de los cuales circula el agua. Dentro de estos órganos, el agua se mueve a través de los tejidos por dos vías: apoplástica (por las paredes de las células y los espacios intercelulares) y simplástica (por adentro de las células). Identificamos con su nombre a diferentes fases del movimiento de agua, absorción (suelo-raíces), transporte (raíces - hojas) y transpiración (hojas - atmósfera), así como a la precipitación la infiltración y la percolación. Esta discusión nos llevó a considerar con algo de detalle la definición de suelo así como su origen y sus características físicas (componentes, secuencia de horizontes, textura y estructura). Vimos que existen suelos con diferente textura, es decir con diferentes proporciones de arcilla, limo y arena en su fracción mineral. Los suelos con textura arcillosa tienen una alta proporción de poros pequeños y los suelos con textura arenosa tienen, en comparación, alta proporción de poros grandes.
En relación con la segunda cuestión (¿cómo?), vimos que el agua se mueve en nuestro sistema siguiendo gradientes de Potencial Hídrico. Vimos que el agua puede estar retenida o impulsada por fuerzas de diferente origen y definimos al Potencial Hídrico como la presión que habría que ejercer para llevarla desde dónde se encuentra hasta un estado de referencia. Examinamos entonces en detalle los diferentes tipos de fuerza que componen el Potencial Hídrico.
Finalmente definimos al Potencial Hídrico como la sumatoria de los 4 componentes.
En relación con la tercera cuestión (¿en qué unidades?), vimos que el flujo de absorción, transporte y transpiración se puede medir en unidades tales como g de agua/planta.día o g de agua/m².día, etc. Además vimos que el Potencial Hídrico y sus componentes se miden en unidades de presión tales como atmósferas o megaPascales (10atm = 1 MPa).
En relación con la última cuestión (¿cuáles son los controles?), primero propusimos que el flujo de agua entre dos puntos es proporcional a la diferencia de Potencial Hídrico entre ellos. Luego vimos cómo puede variar esta diferencia entre el suelo, las hojas y la atmósfera que está en contacto inmediato con las hojas. Vimos que la temperatura, la humedad relativa y la velocidad del viento afectan a el Potencial Hídrico de la atmósfera que rodea a las hojas. El flujo transpiratorio es proporcional a la diferencia entre ese Potencial y el Potencial Hídrico del mesófilo. Además, es inversamente proporcional a la resistencia que aumenta a medida que se cierran los estomas. Finalmente vimos que el Potencial Hídrico del suelo varía con su contenido de agua; es cero en el suelo SATURADO y disminuye, hacíendose cada vez más negativo, a medida que el suelo se seca. La curva que describe la relación entre el Contenido de Agua (en % o en litros de agua/m3 de suelo) y el Potencial Hídrico del suelo (en MPa o en atm) se llama Curva de Retención Hídrica. Este tipo de curva permite calcular la cantidad de AGUA UTIL, que es aquella retenida por fuerzas capilares que las plantas alcanzan a compensar. El agua no retenida por fuerzas capilares es el AGUA GRAVITATORIA que las plantas no aprovechan porque percola. El agua que está en poros muy pequeños no es útil para las plantas porque está tan fuertemente retenida (su Potencial Hídrico es tan bajo) que estas no pueden absorberla. Se denomina CAPACIDAD DE CAMPO al contenido de agua que tiene un suelo luego de una lluvia o riego abundante y cuando ya percoló el agua gravitatoria. Típicamente, en este momento, el Potencial Hídrico del Suelo de de -0.015 MPa. Se denomina PUNTO DE MARCHITEZ PERMANENTE al contenido de agua que tiene un suelo cuando las plantas han extraido toda el agua útil. Comunmente, en ese momento, el Potencial Hídrico del suelo es aproximadamente -1,5 MPa.
En medio de la discusión, a raiz de una pregunta de Fernando, mencionamos muy brevemente que la energía necesaria para generar las diferencias de Potencial Hídrico a lo largo del camino del agua desde el suelo hasta la atmósfera a través de la planta proviene del sol. Es la energía de la radiación solar absorbida por las hojas y transformada en calor latente de vaporización del agua que está en el apoplasto del mesófilo. Esta evaporación disminuye el Potencial Hídrico en los microcapilares de las paredes celulares y genera toda la cadena de tensiones que se transmite hasta el agua que está en los poros del suelo.
Discutimos entre todos los problemas 6 y 7
Analizamos una serie de características y controles del clima que resultan importantes para la productividad de los ecosistemas
Primero examinamos los efectos que la forma de la tierra y la inclinación de su eje de rotación respecto del plano de translación tienen sobre la intensidad de la radiación incidente y sobre la duración del día (los dos componentes de la cantidad total de radiación que incide por día). Vimos que, debido a la curvatura de la tierra, la radiación solar que incide por unidad de superficie terrestre es en términos generales menor cerca de los polos que cerca de los trópicos. Del mismo modo que antes calculamos la radiación incidente sobre una hoja inclinada, ahora podemos calcular la radiación incidente sobre el suelo utilizando el ángulo de incidencia de la luz. La inclinación del eje de rotación de la tierra determina, a su vez, que la duración del día y el ángulo de incidencia de los rayos solares varíen en cada latitud a lo largo del año. Definimos como ECUADOR SOLAR a la latitud en la cual los rayos solares inciden al mediodía en forma perpendicular a la superficie. Vimos que el ecuador solar oscila anualmente entre los trópicos de Cancer y Capricornio. Durante la primavera del hemisferio sur se desplaza desde el Ecuador al trópico de Capricornio. durante el verano del hemisferio sur del trópico de Capricornio al Ecuador, durante el otoño del hemisferio sur del Ecuador al trópico de Cancer y durante el invierno del hemisferio sur se desplaza desde el trópico de Cancer hacia el Ecuador.
Para completar el análisis de la heterogeneidad en la incidencia de radiación vimos que localmente, el ángulo de incidencia varía con las pendientes.
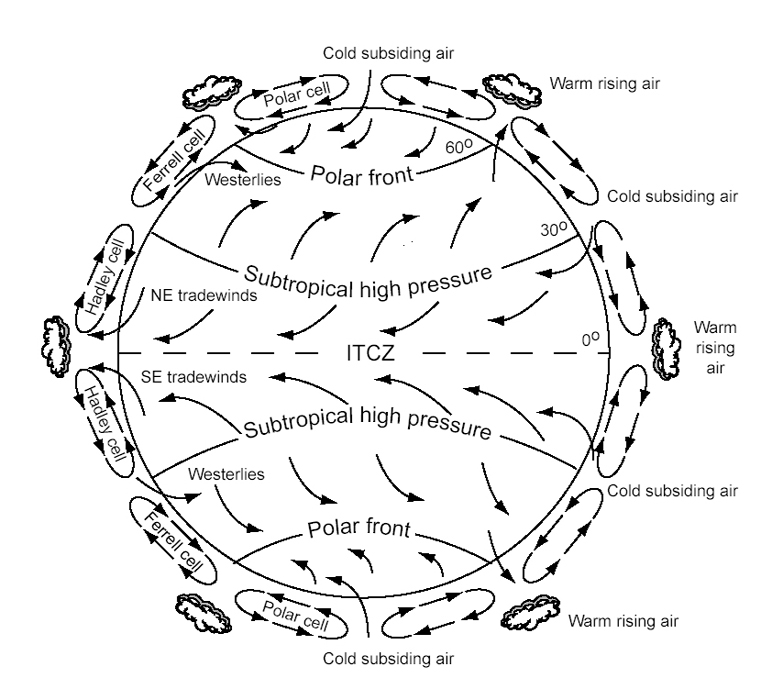
Luego nos concentramos en la distribución de calor y de lluvias. Primero explicamos que mientras la Tierra se calienta porque absorbe radiación solar, la atmósfera se calienta porque absorbe radiación emitida por la Tierra. La atmósfera se calienta mucho cerca del ecuador solar, donde la Tierra se calienta más. Como el aire caliente es más liviano que el aire frío, el calentamiento diferencial de distintas partes de la tierra genera diferencias de presión atmosférica que producen los vientos. Vimos que la circulación del aire en la atmósfera tiene patrones característicos denominados Células de Hadley, Ferrell y Polar. En la célula de Hadley, los vientos predominantes cercanos a la superficie son los Alisios que soplan de este a oeste. En cambio, en la célula de Ferrell los vientos predominantes son los Occidentales que soplan de oeste a este.
Además, vimos que en los centros de baja presión el aire está caliente y asciende. Como consecuencia del ascenso, el aire se enfría, su capacidad de contener humedad disminuye y cuando se enfría por debajo de su punto de rocío, parte del agua que contiene se condensa y llueve. Esto explica la presencia de climas muy lluviosos en la denominada CONVERGENCIA INTERTROPICAL, donde se ponen en contacto las células de Hadley del Hemisferio norte y sur. En contraste, en los centros de alta presión, el aire frio baja, se calienta, no pierde agua y por lo tanto no llueve. Esto explica que cerca de los 30° de latitud, donde se encuentran las células de Hadley y Ferrell formando un cinturón de alta presión aparezcan los principales desiertos del mundo. Para entender el fundamento de estos fenómenos, examinamos con cierto detalle la CURVA DE PRESION PARCIAL DE VAPOR DEL AIRE SATURADO y definimos el PUNTO DE ROCIO y la HUMEDAD RELATIVA. Vimos también que el ascenso y enfriamiento del aire, con las lluvias consecuentes, ocurre también donde los vientos se encuentran con cordones montañosos. Las lluvias que se producen en este caso son denominadas LLUVIAS OROGRAFICAS. El aire que desciende del otro lado de las montañas se calienta por encima de su punto de rocío y no produce lluvias. Esto determina la existencia de zonas áridas en las denominadas SOMBRAS DE LLUVIA.
Finalmente examinamos como las oscilaciones del ecuador solar determinan 3 patrones climáticos definidos: el clima ECUATORIAL, con picos de lluvia en primavera y otoño, cuando el ecuador solar pasa por el ecuador, el clima MONZONICO, con picos de lluvia en verano, cuando el ecuador solar llega al trópico del hemisferio correspondiente y el clima MEDITERRANEO, con picos de lluvia invernales, cuando el ecuador solar alcanza el trópico del hemisferio opuesto y el cinturón de alta presión del hemisferio correspondiente se acerca al ecuador.
En la segunda parte de la clase nos dedicamos a resolver los problemas 9 a 15 para repasar los diferentes temas desarrollados hasta el momento.
En esta clase nos introducimos en la cuestión del flujo de energía en los ecosistemas.
En primer término describimos la estructura de un ecosistema tomando como ejemplo un agroecosistema típico de la región pampeana. Entre todos, hicimos una lista de sus componentes bióticos y abióticos y luego un esquema pictórico de la distribución de estos componentes en el espacio. En un momento, a raiz de la diferencia entre los animales domésticos y silvestres, nuestra discusión llevó naturalmente al concepto de subsidio de energía, la energía necesaria para un trabajo que el hombre efectúa con el objeto de manejar o modular el flujo de energía de un ecosistema. Prometimos analizar ese concepto en más detalle más adelante. En relación con los componentes bióticos del ecosistema, primero distinguimos entre vegetales, animales, hongos, etc. Luego, a partir de la noción de que estos organismos están vinculados por relaciones tróficas, vimos que pueden ser clasificados en los denominados NIVELES TROFICOS (productores, consumidores primarios, consumidores secundarios y detritivoros y descomponedores).
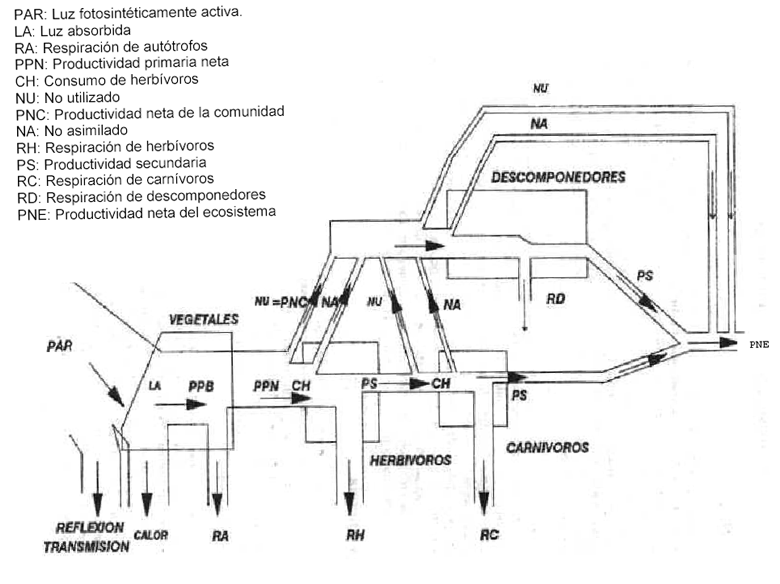
Utilizando el concepto de niveles tróficos, hicimos un esquema sencillo de las posibles tranferencias de energía entre dichos niveles. Vimos que la energía está contenida en el ecosistema en compartimentos correspondientes a la BIOMASA de cada uno de los niveles tróficos y a la NECROMASA. Las transferencias de energía entre estos compartimentos siguen reglas fijas que resumimos en el esquema. Además, nuestro esquema nos permitió visualizar balance de energía del ecosistema, la diferencia entre toda la energía captada por la fotosíntesis de los productors y toda la que sale debido a la respiración de todos los niveles tróficos. Cuando este balance es positivo, su valor es denominado PRODUCTIVIDAD NETA DEL ECOSISTEMA. Finalmente, propusimos unidades de medida apropiadas para la energía contenida en la biomasa de los diferentes niveles tróficos y en la necromasa, las cajas del esquema (p.ej. cal/m²), y para las tasas de transferencia de energía entre estos compartimentos, las flechas de nuestro esquema (p.ej. cal/s.m² o W/m²).
Luego examinamos con más detalle el flujo de energía a través de un nivel trófico cualquiera. Vimos que de toda la energía consumida por este nivel trófico en una unidad de tiempo y superficie, una parte es asimilada y otra parte se pierde. Salvo para los productores, la parte que no es asimilada es transferida a la necromasa (¿Cuál es el destino de la energía no asimilada por los productores?). La tasa de asimilación de energía es la PRODUCTIVIDAD BRUTA del nivel trófico. Una parte de la energía asimilada es disipada por la RESPIRACION y el resto es acumulado en la biomasa. La tasa de acumulación de energía en la biomasa es la PRODUCTIVIDAD NETA del nivel trófico. Una parte de la biomasa acumulada como productividad neta puede permanecer como tal (aumento del tamaño o del número de individuos en el nivel trófico), otra parte es consumida por el nivel trófico siguiente y otra parte muere y se transforma en necromasa. Finalmente distribuímos un esquema del flujo de energía donde están detallados estos diferentes flujos. En ese esquema, la tasa en la cual la energía consumida no asimilada se transforma en necromasa es denominada ENERGIA NO ASIMILADA y la tasa de transferencia de energía de la biomasa a la necromasa por muerte es denominada ENERGIA NO UTILIZADA.
Utilizamos esta clase para resolver y discutir en detalle los problemas 21, 22 y 23. Estos problemas están dirigidos a ejercitar el uso del modelo de flujo de energía en el ecosistema. En particular enfatizamos:
Comenzamos esta clase discutiendo la idea de una pirárámide de energía en el ecosistema. Esta idea se asocia con el hecho de que los niveles tróficos superiores reciben menos energía que los inferiores porque la respiración de cada nivel trófico disipa en forma de calor una parte de la energía que asimila. Una consecuencia de esto es que comúnmente la biomasa y la cantidad de energía contenida en un momento dado por cada nivel trófico disminuye a medida que se pasa de productores a consumidores primarios, consumidores secundarios, etc. Con ayuda de Matías pudimos imaginar una pirámide invertida que representaba las pérdidas de energía por respiración a medida que se avanza en la cadena trófica. Esta discusión anterior nos llevó a pensar otra vez en la productividad neta del ecosistema, que es la diferencia entre la productividad primaria bruta y la suma de las tasas de liberación de energía por respiración de todos los niveles tróficos.
Luego repasamos la definición de productividad primaria neta y razonamos cómo se puede obtener un valor aproximado de la (PPN) a partir de datos de la cantidad de biomasa y necromasa vegetal en fechas sucesivas. Para este razonamiento nos ayudamos con un esquema apropiado del flujo de energía.
Si en un área se impide la entrada de los herbívoros la tasa de acumulación de biomasa vegetal es,
dProd = PPN + NUh,
y la tasa de acumulación de necromasa vegetal (broza) es,
dNecomasa vegetal = NUh - CvD
dónde CvD es la tasa de consumo de detritos vegetales por organismos detritivoros y descomponedores (medida en unidades de energía por unidad
de tiempo y por unidad de superficie). A partir de estas fórmulas se deduce que
PPN = dProd + dNecromasa vegetal + CvD.
En los ecosistemas dónde CvD es relativamente bajo, como es frecuente en los partizales, se puede aproximar la PPN como
PPN = dProd + dNecromasa vegetal.
Después de un intervalo, pasamos a discutir los controles de la PPN. Hicimos una lista de los controles próximos o cercanos de la
PPN que nos resultaban conocidos a partir del desarrollo del tema Fotosíntesis que hicimos en la primera parte del curso:
- agua
- luz
- dióxido de carbono
- nutrientes minerales como el nitrógeno y el fósforo
- temperatura
- tipo de fotosíntesis (C3, C4, CAM).
Luego vimos que estos controles próximos varían en función de factores de acción mediata:
- lluvias
- capacidad de retención hídrica de los suelos
- fertilidad química de los suelos
- latitud
- estructura de la vegetación
- intervenciones humanas tales como riego, fertilización, etc.
Para discutir las respuestas de la PPN a estos factores examinamos una serie de figuras con ejemplos.
Finalmente nos concentramos en las intervenciones humanas que se hacen con el fin de modificar la PPN u otros componentes del flujo de energía. Vimos que se trata de SUBSIDIOS DE ENERGIA porque su aplicación requiere un gasto de energía. En particular examinamos a la aplicación de fertilizante y vimos que para realizarla hay que gastar energía en la producción del fertilizante, en su transporte y en su distribución. Del mismo modo, cualquiera de las intervenciones habituales en los agroecosistemas requiere un gasto de eneregía y es en este sentido que constituye un subsidio. Vimos que pocas veces los subsidios de energía constituyen aportes directos al flujo de energía entre niveles tróficos (como por ejemplo cuando se importa forraje). Normalmente, en realidad, los subsidios de energía constituyen el gasto energético necesario para canalizar la energía de un flujo a otro (por ejemplo canalizar la PPN hacia Consumo de Herbívoros en lugar de hacia No Utilizado).
En esta clase examinamos la circulación del agua, el carbono y el nitrógeno en los ecosistemas.
Comenzamos repasando los controles próximos de la PPN que habíamos enumerado en la clase anterior. En la lista de controles destacamos al agua, el dióxido de carbono y a los nutrientes minerales como el nitrógeno y el fósforo. Recordamos de qué modo actúan sobre la fotosíntesis y cómo son las figuras que muestran su relación con la PPN.
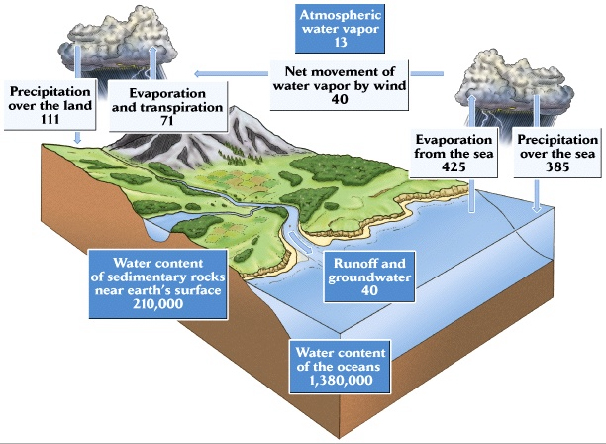
A continuación desarrollamos en detalle el modelo del ciclo grobal del agua. Distinguimos los COMPARTIMIENTOS, cuyo estado se mide en gramos de agua (con magnitudes del orden de 1018 gramos), de los FLUJOS, medidos en gramos/año (con magnitudes del orden de 1018 gramos/año). Los ríos nos dieron la oportunidad de pensar mejor la diferencia entre comparimiento y flujo. Vimos que, junto con los lagos, constituyen un compartimiento que contiene unos 100x1018 gramos de agua y que, al mismo tiempo, determinan un flujo hacia los mares de unos 40x1018 gramos de agua por año. Luego, calculamos el TIEMPO DE RESIDENCIA del agua en un compartimiento como el cociente entre su contenido de agua y la suma de los flujos de salida del compartimiento en cuestión. A partir de este cálculo vimos que mientras el tiempo de residencia del agua en la atmósfera es de menos de dos semanas, su tiempo de residencia en los continentes (incluyendo el agua de los lagos y ríos, el suelo y los organismos) es de casi 2 años y en los mares es de más de 3000 años. Finalmente formulamos un modelo mucho más detallado para la circulación local del agua en la escala de un potrero.
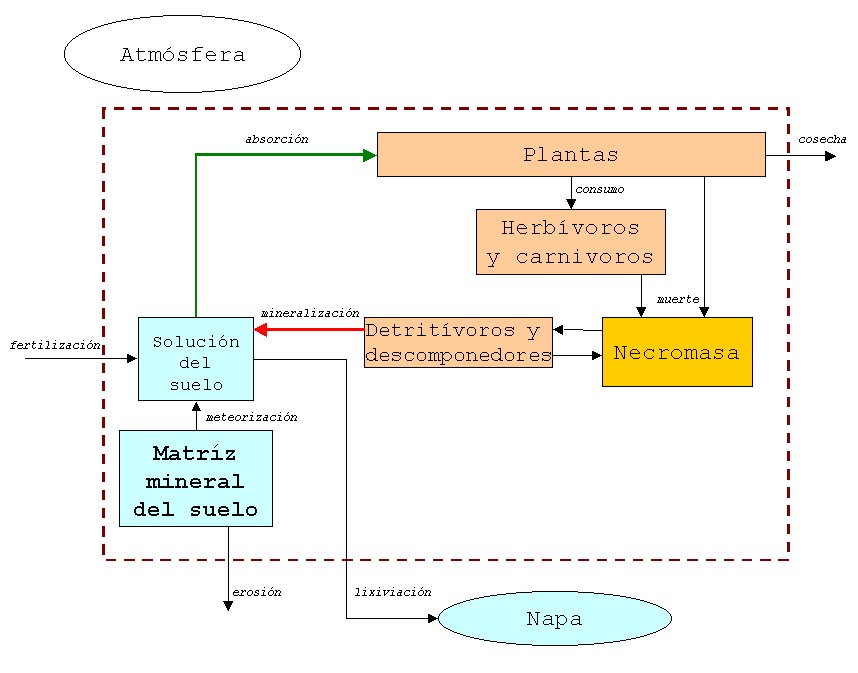
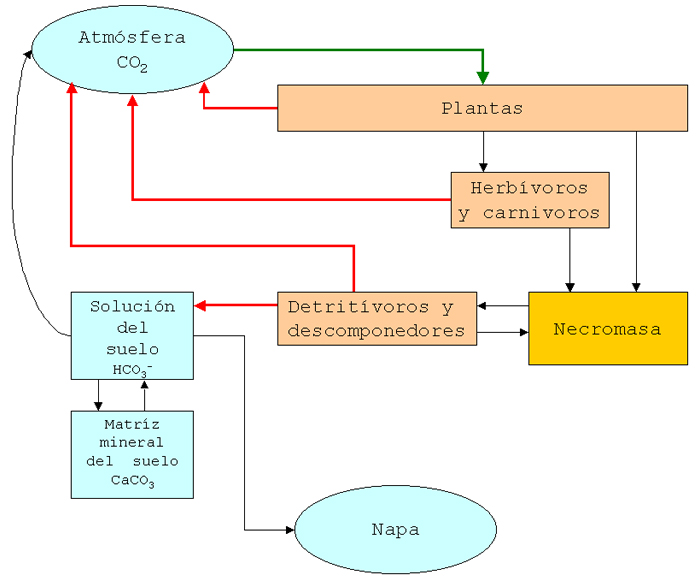
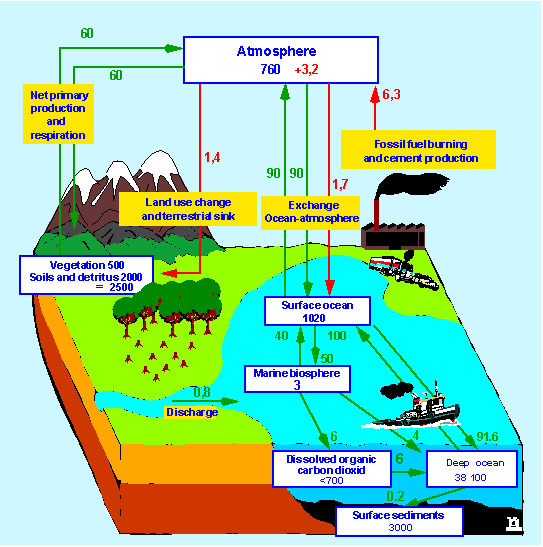
Luego desarrollamos un modelo de la circulación del carbono en la escala de ecosistema local utilizando como base el problema 30. Para ello, primero identificamos las formas en que el carbono está contenido en los compartimientos Atmósfera, Plantas, Consumidores, Necromasa, Descomponedores, Solución del Suelo, Matriz mineral del Suelo y Napa, y luego detallamos cuáles flujos de carbono ocurren entre estos compartimientos. En relación con las formas en que se encuentra el carbono en el ecosistema distinguimos especialemnte las formas orgánicas e inorgánicas. En relación con los flujos, destacamos que el proceso asimilador de carbono (transformación de carbono inorgánico en orgánico) es la fotosíntesis, que transfiere carbono de la atmósfera a las plantas, y el proceso desasimilador es la respiración de las plantas, consumidores y descomponedores que transfiere dióxido de carbono a la atmósfera o a la solución del suelo.
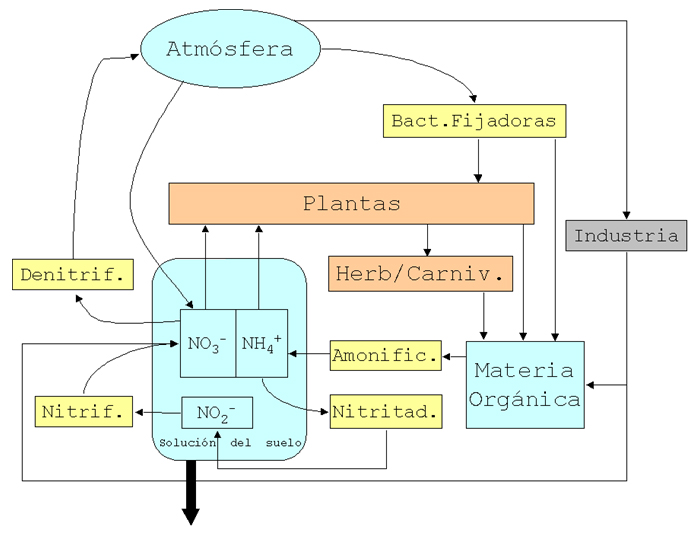
Finalmente usamos el problema 30 para examinar la circulación del nitrógeno en la escala de ecosistema local. En este caso, también comenzamos por identificar las formas en que se encuentra este elemento en los diferentes compartimientos para luego detallar los flujos entre ellos. El principal reservorio de nitrógeno es la atmósfera. Los flujos naturales desde la atmósfera hacia los restantes compartimientos son la deposición de formas de nitrógeno inorgánico como amonio o nitrato disueltos en el agua de lluvia y la fijación biológica, un proceso asimilador que incorpora nitrógeno atmosférico a compuestos orgánicos producidos por ciertas bacterias o algas fijadoras. Para incluír este último proceso en nuestro modelo, agregamos un compartimiento denominado "fijadores" y los flujos correspondientes. Destacamos que la principal fuente de nitrógeno para las plantas es el que se encuentra en forma de amonio o nitrato en la solución del suelo. Esto nos llevó a detallar en el modelo los procesos que dereminan la disponibilidad de estas formas de nitrógeno. Para ello, tuvimos que agregar los compartimientos correspondientes a las "bacterias nitrificadoras", que transforman el amonio que resulta de la descomposición de las moléculas orgánicas nitrogenadas en nitratos, y a las bacterias "denitrificadoras", que transforman los nitratos en nitrógeno molecular. Mencionamos a la difusión de este nitrógeno molecular hacia la atmósfera como uno de los flujos de naturale de salida de nitrógeno del ecosistema. Además mencionamos la salida por lixiviación de nitrógeno de la solución del suelo hacia la napa (En realidad también sale nitrógeno como amoníaco y óxidos de nitrógeno volátiles). Para finalizar mencionamos algunos flujos de entrada y salida de nitrógeno de orígen antrópico: la entrada por fertilización y la salida por extracción de los productos de la cosecha. Ver Diagrama elaborado en clase.
Al comienzo de esta clase examinamos la circulación del fósforo en los ecosistemas, tema que nos había quedado pendiente. Juntos elaboramos un diagrama que nos sirvió de modelo general para la circulación de los nutrientes de ciclo sedimentario. Sobre ese diagrama, llamamos la atención sobre el hecho de que, en la escala de tiempo ecológica, este tipo de nutrientes, que tiene como su principal fuente el sustrato mineral, tiende a perderse de los ecosistemas terrestres.
Luego discutimos el proceso de cambio global apoyándonos principalmente en cinco modelos conceptuales que analizamos antes en en curso:
- El ciclo del carbono,
a partir del cual analizamos un esquema de su circulación
global
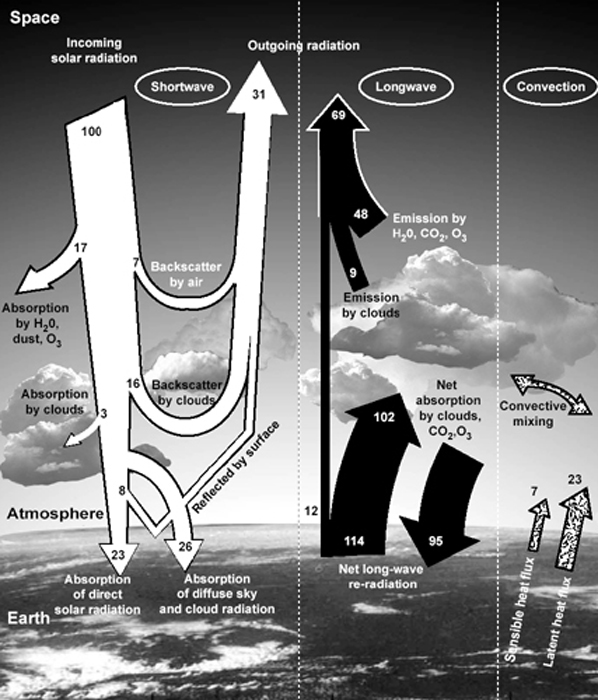
- El balance de radiación de la Tierra.
- La circulación global de la atmósfera.
- El flujo de energía en los ecosistemas.
- La circulación del nitrógeno y su relación con el flujo de energía.
Para finalizar, introdujimos brevemente en concepto de SERVICIOS DE LOS ECOSISTEMAS y diferenciamos los que tienen valor de mercado de los que no lo tienen y, por tal razón el costo de su pérdida es una externalidad para quienes explotan los ecosistemas.
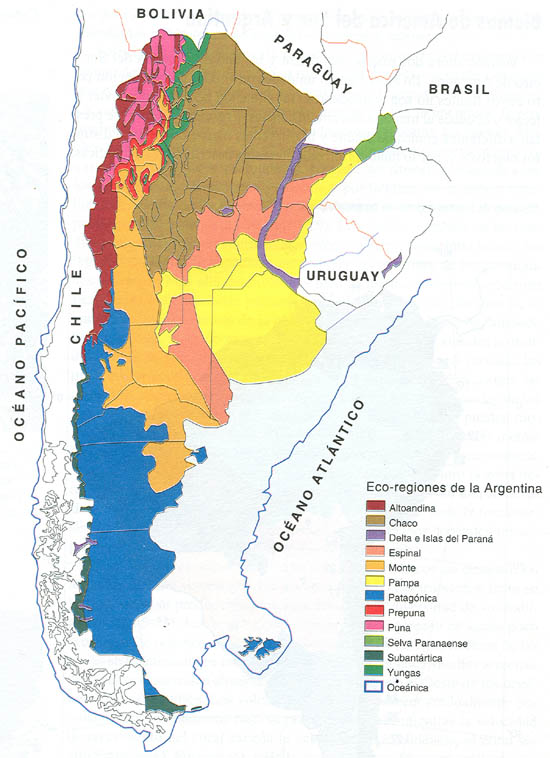
A partir de una serie de fotos que ilustran estos biomas y tipos de vegetación de la Argentina caracterizamos su distribución en diferentes regiones ecológicas, e identificamos características de su estructura y de su funcionamiento en las que difieren. En principio, pusimos especial énfasis en las características del clima con el cual están asociados los diferentes tipos de vegetación pero a partir de varios ejemplos mostramos que existen otros factores que influyen tanto sobre la estructura y sobre el funcionamiento de los ecosistemas y que determinan su heterogeneidad en el espacio. Para finalizar, elaboramos un modelo conceptual de los factores que controlan dicha heterogeneidad en la estructura y funcionamiento de los ecosistemas. En este modelo incluimos al clima, el suelo, la biota presente, las interacciones entre organismos, los disturbios y el uso antrópico.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}